微生物构成的氮循环网络
The microbial nitrogen-cycling network
翻译:翟志文 中科院遗传发育所
责编:刘永鑫 中科院遗传发育所
Nature Reviews Microbiology, [31.851]
https://doi.org/10.1038/nrmicro.2018.9
Published: 05 February 2018
第一作者:Marcel M. M. Kuypers
通讯作者:Marcel M. M. Kuypers mkuypers@mpi-bremen.de
合作作者:Hannah K. Marchant and Boran Kartal
作者单位: 德国,柏林,马普海洋微生物研究所
主编按:能发表在30分以上的综述,都是阅读了500-1000篇文献,大牛花了整年的时间,总结的教科书级别的学习材料,相关领域的学生有必要多读几遍。目前此文发表仅20个月,Google统计引用151次。而且氮是任何生命的重要元素,无论你研究的对象是动物、植物、微生物或环境,相信有本文氮循环的背景知识,定会让你文章的结果和讨论增色不少。
一作和兼通讯作者Marcel Kuypers,目前发表本领域是相关文章176篇,其中包括Nautre、Science、PNAS等众多顶级期刊,引用达1.8万次,H指数高达67。
就看这专业背景,读不读你自己选!
大纲
Outline
摘要
Abstract
氮是所有生物的重要组成部分,也是限制我们星球上所有生命体的主要营养成分。目前为止,可自由获得氮的主要来源是大气中的氮气,但是大部分生物是依赖生物可利用的氮形式来生长,比如铵盐和硝酸盐。而这些基质的可利用性是依赖于不同的氮转化反应,这些氮转化反应是由具有多种多样新陈代谢功能的微生物构成的复杂网络而实现的。本综述在目前获得的新研究和新发现的背景下,对当前由微生物参与的氮转化过程进行描述,包括这些微生物构成的氮转化反应,参与氮转化的微生物以及它们的生理和环境功能,也对那些并未发现但可能会发生的反应进行了描述。此外,我们对由氮转化微生物之间的相互作用而构成的复杂网络和其对全球生物化学氮循环的影响进行了讨论。
Nitrogen is an essential component of all living organisms and the main nutrient limiting life on our planet. By far, the largest inventory of freely accessible nitrogen is atmospheric dinitrogen, but most organisms rely on more bioavailable forms of nitrogen, such as ammonium and nitrate, for growth. The availability of these substrates depends on diverse nitrogen-transforming reactions that are carried out by complex networks of metabolically versatile microorganisms. In this Review, we summarize our current understanding of the microbial nitrogen-cycling network, including novel processes, their underlying biochemical pathways, the involved microorganisms, their environmental importance and industrial applications.
亮点
Key points
氮是所有生物的基本组成部分,也是限制地球生命的主要营养物质。
其可用性取决于微生物进行的多种氮转化反应。
Nitrogen is an essential component of all living organisms and the main nutrient limiting life on our planet. Its availability depends on diverse nitrogen-transforming reactions that are carried out by microorganisms.
氮转化微生物在代谢上是多功能的,这使得它们的分类仅仅局限于硝化器、反硝化器和类似的类别。
Nitrogen-transforming microorganisms are metabolically versatile, rendering their classification as mere nitrifiers, denitrifiers and similar classes inadequate.
经典的氮循环是不可能存在的,它由不同的过程以有序的方式相互遵循组成。
在自然界中,微生物形成了连接氮转化反应的复杂网络。
The classical nitrogen cycle consisting of distinct processes that follow each other in an orderly fashion does not exist. In nature, microorganisms form complex networks that link nitrogen-transforming reactions.
微生物氮转化网络会减弱和加剧由人类引起的全球变化。
它们产生和消耗强大的温室气体一氧化二氮(氧化亚氮,N2O),导致水生系统富营养化,同时从废水中去除氮。
Microbial nitrogen-transforming networks both attenuate and exacerbate human-induced global change. They produce and consume the powerful greenhouse gas nitrous oxide, lead to eutrophication of aquatic systems and, at the same time, remove nitrogen from wastewater.
还有许多未发现的氮转化反应是热力学上可行的。
催化这些反应的微生物以及相关的生化途径等着我们去发现。
There are still many undiscovered nitrogen-transforming reactions that are thermodynamically feasible. The microorganisms catalysing these reactions and the involved biochemical pathways are waiting to be discovered.
名词解析
还原剂:
氧化还原反应中的给电子供体化合物。
Reductants:The electron-donating compounds in a redox reaction.
产氧光养生物:
从光中获取能量并利用水作为电子供体,形成分子氧和糖作为产物的生物。
Oxygenic phototrophs: Organisms that obtain energy from light and use water as the electron donor, forming molecular oxygen and sugar as products.
细菌细胞:
动物体内含有内共生细菌的特殊细胞。
Bacteriocytes: Special cells in animals that contain endosymbiotic bacteria
奇古菌门:
含有氨氧化古细菌的门。
Thaumarchaeota: The phylum that contains the ammonia-oxidizing archaea.
嗜酸性:
生物体在酸性环境中生长的倾向(pH < 6)。
Acidophilic:The propensity of organisms to grow in acidic environments (pH <6).
甲烷:
转化生物氧化甲烷以保存能量。
Methanotrophs: Organisms that oxidize methane to conserve energy.
NC10:
以澳大利亚纳拉伯洞穴命名的一个候选细菌门,包含“甲基假丝酵母oxyfera”,这是第一个发现的进行甲烷氧化和氧反硝化的有机体。
NC10:A candidate bacterial phylum, named after the Nullarbor Caves in Australia, that contains ‘Candidatus Methylomirabilis oxyfera’, which is the first organism discovered that performs methane oxidation coupled to oxygenic denitrification.
反能:
需要能量输入的反应。
Endergonic: A reaction that requires energy input.
疣菌门:
一种细菌门,仅有少数被描述的物种,其中一些似乎在甲烷循环中很重要。
Verrucomicrobia: A bacterial phylum with only a few described species, some of which appear to be important in the methane cycle.
缺氧光致养生物:
这些微生物从光中获取能量,并用硫化氢等化合物代替水作为电子供体,因此不产生分子氧。
Anoxygenic phototrophs: These microorganisms obtain energy from light and use compounds such as hydrogen sulfide instead of water as an electron donor and thus do not produce molecular oxygen.
富营养化:
营养物质的增加,导致藻类或蓝藻过度生长。
Eutrophication: An increased input of nutrients that leads to excessive growth of algae or cyanobacteria.
质子动力:
质子位错在细胞膜两侧产生电荷和酸碱度的差异,从而产生一个电化学电位,用于节能。
Proton motive force: Proton dislocation creates a difference of charge and pH between two sides of a cell membrane and thereby generates an electrochemical potential, which is used for energy conservation.
厌氧污泥硝化器:
在废水处理过程中产生的多余微生物生物量(污泥)厌氧转化为二氧化碳、甲烷、铵和还原硫化合物的生物反应器。
Anaerobic sludge digesters: Bioreactors in which excess microbial biomass (sludge) produced during wastewater treatment is anaerobically converted to carbon dioxide, methane, ammonium and reduced sulfur compounds.
初级亚硝酸盐最大值:
在透光区底部亚硝酸盐浓度的峰值。
Primary nitrite maxima: The peak in nitrite concentrations at the base of the euphotic zone.
一氧化氮歧化:
两个一氧化氮分子歧化成一个分子氧和一个分子二氮气体。
Nitric oxide dismutation: Two molecules of nitric oxide are disproportionated into one molecule of molecular oxygen and one molecule of dinitrogen gas.
化合作用:
两种反应物中含有同一种元素,氧化状态不同,反应生成一种氧化状态单一的产物的化学反应。
Comproportionation: A chemical reaction in which two reactants containing the same element with a different oxidation state react to create a product with a single oxidation state.
厌氧氨氧化菌:
在厌氧氨氧化菌(anammox)中发现的一种细菌细胞器,是唯一已知的原核膜结合结构,在细胞分裂时被均分为子细胞。
Anammoxosome: A bacterial organelle found in anaerobic ammonium oxidizing (anammox) bacteria that is the only known prokaryotic membrane-bound structure that is equally divided into daughter cells upon cell division.
放能:
产生自由能的反应。
Exergonic: A reaction that results in the release of free energy.
歧化反应:
一种化学反应,其中一种反应物被分成两种,含有相同的元素,具有不同的氧化状态,一种被氧化,另一种比反应物还原。
Disproportionation: A chemical reaction in which a reactant is split into two species containing the same element with different oxidation states, one more oxidized and the other more reduced than the reactant.
背景和内容概述
氮是所有生物体不可缺少的组成元素,也是合成蛋白质和核酸等关键细胞化合物的必要物质。大气中的氮气是可自由获得氮的最大库存,从生物学上来讲,它具有高度多样的可利用性,然而能够固氮的细菌和古生物菌是很少的。而其他生物体却必须依靠如铵盐和硝酸盐这种性质活泼的含氮物质来提供生长,可这些生物可利用的氮在许多环境中是罕见的。这些限制性营养盐的可获得性主要是通过由微生物参与的、能够改变氮氧化状态的反应来控制。
由于工业氮肥的高投入量,导致人类活动对生物可利用氮的数量产生了极大的影响,人类生产的50%的食物需要依靠工业氮肥,这种氮肥的使用和豆科植物的种植几乎使向陆地和海洋生态系统中的氮输入量增加了一倍。因此为了预测这种氮输入的后果,我们迫切地需要了解由微生物构成的氮转化的基本机制。
由微生物形成的氮转化的过程通常被描述为由六个有序进行的不同反应过程组成的一个循环。这种观点认为一个氮气分子(N2首先通过固氮作用变成氨气,氨气经过同化吸收作用而转变成生物有机氮,有机氮再经过氨化作用转变成铵盐,而铵盐再通过硝化作用被氧化成硝酸盐(NH4+ → NO2- → NO3-),最终再经反硝化作用被还原为一个氮气分子(NO3- → NO2- → NO → N2O → N2)或者经厌氧氨氧化(anaerobic ammonium oxidatio)作用被还原为一个氮气分子(NO2- + NH4+ → N2)。事实上,在氮循环过程并未达到上述所描述的一个平衡状态,反而是与这六个过程中截然不同的氮通量的大小有关(Box 1)。
氮转化微生物一般按其所涉及的六个过程之一进行分类:硝化器进行硝化;反硝化器、反硝化;固氮器、固氮;等等(Nitrogen-transforming microorganisms are generally classified according to one of the six processes they are involved in: nitrifiers carry out nitrification; denitrifiers, denitrification; nitrogen-fixers, nitrogen fixation; and so on)。过去十年来收集的基因数据揭示了参与氮转化的微生物实际上存在巨大的多种多样的新陈代谢功能。现在我们知道很多微生物可以同时发生固氮和脱氮作用。因此,由于它们代谢的多样性,几乎不可能再根据氮转化过程中经典的六个过程来划分这些微生物。故在本文中,我们将使用过程的名字,比如反硝化作用denitrification和硝化作用nitrification,,而避免对微生物进行相应的分类。这篇综述集中在氧化还原反应,转化氮化合物,生化途径,以及负责的酶(图1)和微生物。
根据我们目前的理解,微生物可通过14个离散型的氧化还原反应改变从-3价态到+5价态的含氮化合物(图1),有机氮和氨气之间的相互转化中氧化还原状态不变。氮转化有关的酶经常在不同的微生物中被发现,而且这里包括的很多酶还是最近才被鉴定出来的。在过去的十年里,有四种新的反应被研究学者发现,它们分别是:羟胺氧化为一氧化氮的反应(hydroxylamine oxidation to nitric oxide,图1,反应7);一氧化通过歧化作用转变成氮气和氧气的反应(nitric oxide dismutation to dinitrogen gas and oxygen,图1,反应9);联氨的合成反应(hydrazine synthesis,图1,反应13);联氨氧化成氮气的反应(hydrazine oxidation to dinitrogen gas,图1,反应12)。另外,许多新的代谢能力也被发现,比如光氧亚硝酸盐氧化(phototrophic nitrite oxidation)和生成硝酸盐的完全氨氧化(complete ammonia oxidation (comammox))。许多新型的微生物也被鉴定出来,比如氨氧化古生菌(ammonia-oxidizing archaea14),反硝化真核生物有孔虫(denitrifying
eukaryotic foraminifera)和共生异养的固氮蓝藻(symbiotic heterotrophic
nitrogen-fixing cyanobacteria)。
本文中,我们在这些新发现的背景下,对目前由微生物引起的氮转化过程进行理解,并对这些微生物构成的氮转化反应,参与氮转化的微生物以及它们的生理和环境功能进行描述,也对那些并未发现但可能会发生的反应进行了描述。此外,我们对由氮转化微生物之间的相互作用而构成的复杂网络和其对全球生物化学氮循环的影响进行了讨论。
Box 1 | 氮全球生化循环圈:全球的氮库存、氮转化过程、氮通量
Box 1 | Biogeochemical nitrogen cycling: global inventories, processes and fluxes
图Box 1. 氮全球生化循环圈:全球的氮库存、氮转化过程、氮通量
如图Box 1所示,图中灰色方块表示氮储存量,束缚在岩石和沉积物中的最大氮存量是氨(1.8×1010 Tg氮),虽然这种束缚氨经侵蚀后可被生物利用,但是它在整个氮循环生化圈中起的作用是很小的,而陆地中可自由获得的氨的储存量是未知的,海洋中的氨的氮储存量被估计为340 ~ 3600 Tg;全球中可自由获得氮的储存库是大气中的氮气(约为3.9×109 Tg氮), 紧接着是有机氮、硝酸盐、氧化亚氮中氮的储存量;而全球的亚硝酸盐和一氧化氮中的氮储存量可以忽略不计。
在这些氮储存库中形成的全球生化的氮循环主要是因为以下六个不同的氮转化过程:同化作用(Assimilation)、氨化作用(ammonification)、硝化作用(nitrification)、反硝化作用(denitrification)、厌氧氨氧化作用(anammox) 和固氮作用(nitrogen fixation)。我们从可查文献中估计了这些过程中的年度氮通量,并作出了如图Box 1所示的简单的假设,在本图中,我们用每年所含 Tg氮来表示在这些主要含氮的物质之间的氮通量,其中绿色、蓝色和红色方块分别代表每年的陆地氮通量、每年的海洋氮通量以及每年人为制造的氮通量。其中最明确的氮通量包括氮的流失和固定,因为它们是重点研究的对象。如图Box 1所示,虽然这些明确的氮通量相对较小,但它却可以调控这些生物可利用氮的可用性,而这些生物可利用氮在很大程度上可以通过生物碳泵来控制大气中二氧化碳的去除。据目前估计,生物固氮量(300 Tg氮/年)和人工固氮量(125 Tg/年)的总和已经超过了通过厌氧氨氧化和反硝化作用产生的氮气的量(350 Tg氮/年)。并不是所有的经一氧化氮还原产生的氧化亚氮都可以进一步的被还原为氮气,而每年从海洋和陆地环境中释放的氧化亚氮的氮通量分别为4 Tg和12 Tg,虽然与其他氮形式的通量相比,氧化亚氮的氮通量相对较小,但是它对环境影响却很大,因为氧化亚氮是破坏臭氧层的主要气体,而且它还是一个强大的温室气体。
如图Box 1所示,在这些不同的氮转化过程中有着截然不同的氮通量,它们并没有达成一个平衡的氮循环,这和论文及教材中经常描述的一样,而最大的氮通量跟氨气和有机氮的相互转化有关。仅在海洋环境中,每年与氨化作用和氨同化作用有关的氮通量(8800 Tg氮/年)比海洋中氮的流失和获得的氮通量总和(约400 Tg氮/年)高出了一个数量级。另一个巨大的氮通量(约2000 Tg氮/年)跟硝化作用有关,这也正好可以解释为什么在海洋中氨盐的浓度虽然不高,但是海洋氨氧化古生菌却是最丰富的微生物之一。与硝酸盐同化有关的氮通量和硝化作用的氮通量是一个数量级的,即海洋浮游植物每年可减少约2000 Tg的硝酸盐氮。而与之相比,异化硝酸盐还原为氨的氮通量可能会比较小。虽然目前还没有可用的陆地环境氮通量估计的数据,但是陆地植物与海洋植物相比对氮的需求比较低。对于陆地植物来说,平均每固定40个分子的碳才需要1分子的氮,而对于海洋植物来说,平均每固定6.6个分子的碳就需要1分子的氮,所以陆地中与同化作用相关的氮通量大概是海洋中的1/6。所以假设在稳定状态下(即氮的获得和流失相等),我们用海洋中的氮通量除以6来估计陆地环境中硝化作用与氨化作用中的氮通量。
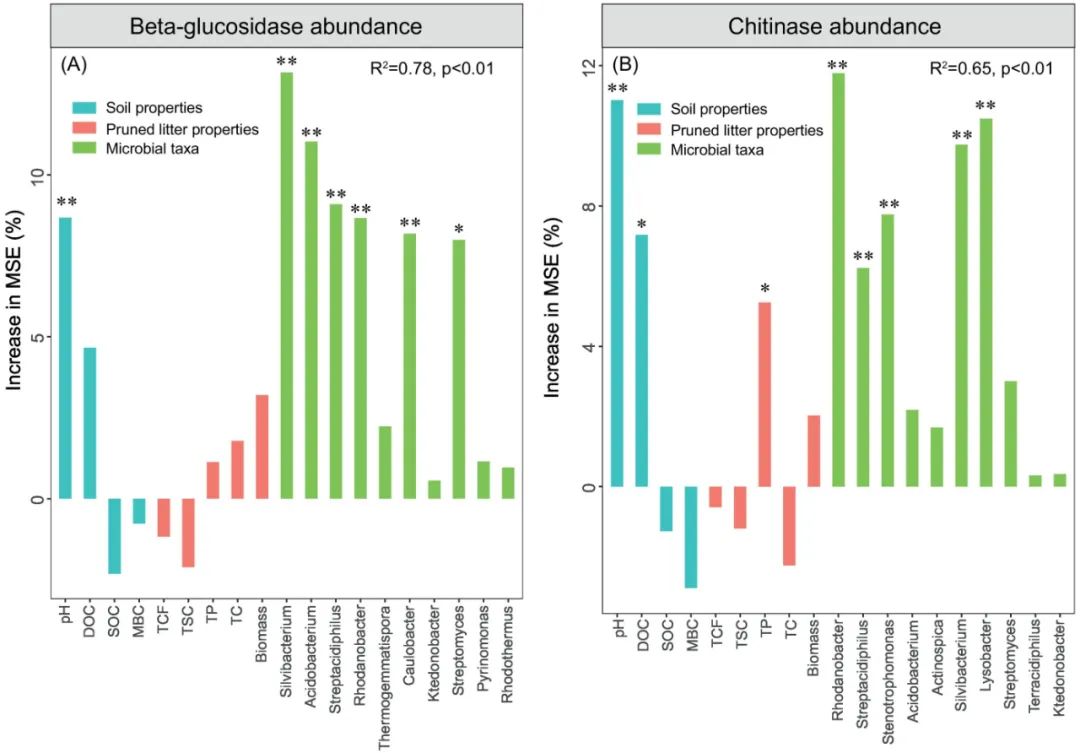
图1 | 微生物转化氮化合物
图1. 微生物构成的氮化合物的转化
如图1所示微生物携带的酶可以执行14个氧化还原反应,在这些氧化还原反应中主要包括8种不同氧化状态下的无机氮(限制酶的中间产物和它们的氧化还原状态没有显示)。氨气与有机氮的相互转化不涉及氮原子氧化还原状态的改变。其中红色的表示还原反应,蓝色的表示氧化反应,绿色的表示歧化反应(disproportionation)和归中反应(comproportionation)。接下来是执行各种反应的酶:同化硝酸盐还原酶(NAS、nasA和nirA);膜结合酶(NAR, narGH);周质异化硝酸还原酶(NAP, napA);亚硝酸盐氧化还原酶(NXR, nxrAB);一氧化氮氧化酶(NOD, hmp);含血红素(cd1-NIR, nirS)和含铜的亚硝酸还原酶(Cu-NIR, nirK);依赖红细胞色素(cNOR, cnorB)、依赖对苯二酚(qNOR, norZ)和含铜的依赖对苯二酚一氧化氮还原酶(CuANOR);依赖NADH细胞色素P450的一氧化氮还原酶(P450NOR, p450nor);黄铜-二铁一氧化氮还原酶(NORvw, norVW);杂交簇蛋白(HCP, hcp);羟胺氧化还原酶(HAO, hao);羟胺氧化酶(HOX, hox);氧化亚氮还原酶(NOS, nosZ);一氧化氮歧化酶(NO-D, norZ);同化亚硝酸盐还原酶(cNIR, nasB和nirB);周质异化细胞色素C亚硝酸盐还原酶(ccNIR, nrfAH);ε-羟胺氧化还原酶(εHAO, haoA);八面体血红素亚硝酸盐还原酶(ONR);八面体血红素连四硫酸盐还原酶(OTR);钼-铁(MoFe, nifHDK)、铁-铁(FeFe, anfHGDK)和钒-铁(VFe, vnfHGDK)固氮酶;联氨脱氢酶(HDH, hdh);联氨合成酶(HZS, hzsCBA);氨单加氧酶(AMO, amoCAB);微粒甲烷单加氧酶(pMMO, pmoCAB);氰酸酯酶(CYN, cynS); 尿素酶(URE, ureABC)。
Fig. 1 | Microbial transformations of nitrogen compounds. Microorganisms carry enzymes that perform 14 redox reactions involving 8 key inorganic nitrogen species of different oxidation states (enzyme-bound intermediates and their redox states are not shown). The interconversion of ammonia and organic nitrogen does not involve a change in the redox state of the nitrogen atom. The reactions involve reduction (red), oxidation (blue) and disproportionation and comproportionation (green). The following enzymes perform the nitrogen transformations: assimilatory nitrate reductase (NAS, nasA and nirA); membrane-bound (NAR, narGH) and periplasmic (NAP, napA) dissimilatory nitrate reductases; nitrite oxidoreductase (NXR, nxrAB); nitric oxide oxidase (NOD, hmp); haem-containing (cd1‑NIR, nirS) and copper-containing (Cu‑NIR, nirK) nitrite reductases; cytochrome c‑dependent (cNOR, cnorB), quinol-dependent (qNOR, norZ) and copper-containing quinol-dependent nitric oxide reductases (CuANOR); NADH-dependent cytochrome P450 nitric oxide reductase (P450NOR, p450nor); flavo-diiron nitric oxide reductase (NORvw, norVW); hybrid cluster protein (HCP, hcp); hydroxylamine oxidoreductase (HAO, hao); hydroxylamine oxidase (HOX; hox); nitrous oxide reductase (NOS, nosZ); nitric oxide dismutase (NO‑D, norZ); assimilatory nitrite reductase (cNIR; nasB and nirB); dissimilatory periplasmic cytochrome c nitrite reductase (ccNIR, nrfAH); ε-hydroxylamine oxidoreductase (εHAO; haoA); octahaem nitrite reductase (ONR); octahaem tetrathionate reductase (OTR); molybdenum-iron (MoFe, nifHDK), iron-iron (FeFe, anfHGDK) and vanadium-iron (VFe, vnfHGDK) nitrogenases; hydrazine dehydrogenase (HDH, hdh); hydrazine synthase (HZS, hzsCBA); ammonia monooxygenase (AMO, amoCAB); particulate methane monooxygenase (pMMO, pmoCAB); cyanase (CYN, cynS); and urease (URE, ureABC).
图2 | 催化氮循环中关键的四种反应的酶
图2 | 催化四个关键氮循环反应的酶。
a | 钼铁(molybdenum-iron,MoFe)固氮酶包含电子转移蛋白(绿色;由nifH编码)和催化酶的α亚单位(洋红;由nifD编码)和β亚单位(紫色;由nifK编码)。nifH用于检测环境中的固氮微生物。铁硫簇介导电子转移到催化中心。电子转移和催化蛋白的结合和解离需要输入ATP。
b | 在厌氧氨氧化细菌库尼尼亚-斯图加滕斯中,电子流过八胺羟胺氧化酶(红色箭头)。属于不同单体的血红素用绿色、蓝色和灰色表示。Haem 4是催化中心。
c | 在膜结合细菌硝酸还原酶(NAR)中,催化二聚体由narG和narH编码,而膜锚由narI编码。narG用于检测环境中的反硝化微生物。硝酸盐还原为亚硝酸盐发生在细胞质,质子转移到周质体。因此,NAR对质子动力有贡献。
d | 在斯图加滕氏杆菌中,hzsA、hzsB和hzsC编码联氨合酶。hzsA、hzsB用于环境中厌氧氨氧化菌的检测。这种酶被认为进行两步反应。开始于γ亚单位(灰色),一氧化氮还原为羟胺,通过底物通道(棕色)转运至α亚单位(绿色)。α亚单位将羟胺与氨水化合为联氨。这两种反应都是由细胞色素c型血红素蛋白催化的。
*氟化ADP-铵;ATP类似物。a部分经david goodsell,doi:10.2210/rcsb_pdb/mom_2002_2(2002年2月)许可改编。b部分最初Maalcke,W.J.等人发表在《生物化学杂志》上八亚甲基氧化还原酶产生生物一氧化氮的结构基础。2014年;289:1228-1242。(参考文献8)。c部分经Elsevier参考文件154的许可进行了修改。d部分经麦克米伦出版有限公司参考文件115的许可进行改编。
Fig. 2 | Enzymes catalysing four key nitrogen-cycling reactions.
a | The molybdenum-iron (MoFe) nitrogenase enzyme contains the electron transfer protein (green; encoded by nifH) and the α-subunit (magenta; encoded by nifD) and β-subunit (purple; encoded by nifK) of the catalytic enzyme. nifH is used to detect nitrogen-fixing microorganisms in the environment. The iron-sulfur clusters mediate electron transfer to the catalytic centre. The association and dissociation of the electron transfer and catalytic proteins require the input of ATP.
b | In the anaerobic ammonium-oxidizing bacterium Kuenenia stuttgartiensis, electrons flow through the haems of the octahaem hydroxylamine oxidase (red arrows). Haems belonging to different monomers are depicted in green, blue and grey. Haem 4 is the catalytic centre.
c | In the membrane-bound bacterial nitrate reductase (NAR), the catalytic dimer is encoded by narG and narH, whereas the membrane anchor is encoded by narI. narG is used to detect denitrifying microorganisms in the environment. Nitrate reduction to nitrite occurs in the cytoplasm, and protons are translocated into the periplasm. Thereby, NAR contributes to the proton motive force.
d | In K. stuttgartiensis, hzsA, hzsB and hzsC encode hydrazine synthase. The former two genes are used to detect anaerobic ammonium-oxidizing bacteria in the environment. This enzyme is proposed to perform a two-step reaction. It starts in the γ-subunit (grey) with the reduction of nitric oxide to hydroxylamine, which is transported through the substrate channel (brown) to the α-subunit (green). The α-subunit comproportionates hydroxylamine with ammonia into hydrazine. Both reactions are catalysed by cytochrome c‑type haem proteins.
*ADP-ammonium fluoride; ATP analogue. Part a is adapted with permission from David Goodsell, doi:10.2210/rcsb_pdb/mom_2002_2 (February 2002). Part b was originally published in the J. Biol Chem. Maalcke, W. J. et al. Structural basis of biological NO generation by octaheme oxidoreductases. 2014; 289: 1228–1242. © the American Society for Biochemistry and Molecular Biology (REF. 8). Part c is adapted with permission from REF. 154, Elsevier. Part d is adapted with permission from REF. 115, Macmillan Publishers Limited.
氮转化反应
Nitrogen-transforming reactions
固氮作用
Nitrogen fixation
大气中的氮气是可以自由获得氮的最大库存,但从生物学上讲,只有微生物携带的金属固氮酶可以将氮气固定为氨(ammonia)。固氮酶普遍分布在细菌和古生菌中,这使它们在生物可利用氮被消耗的环境中存在非常有利的竞争优势。总共有三种类型的固氮酶:铁-铁(FeFe)、钒-铁(VFe)和钼-铁(MoFe)固氮酶,它们有相似的序列、结构和功能特性,但它们的金属辅因子(cofactor)不同,而所有的固氮酶都是有两种物质组成(图2a)。anfDGK,vnfDGK或nifDK分别在活动中心分别编码含铁、钒或钼的固氮酶的催化成分。此外,anfH、vnfH 或 nifH 编码含铁的电子转运蛋白(又称固氮还原酶nitrogenase reductase或铁蛋白iron protein)。NifH是用来检测环境中有固氮作用微生物的标记基因。土壤细菌Azotobacter vinelandii可以编码全部三种类型的固氮菌,然而其它微生物,比如海洋固氮菌 Trichodesmium spp. 只含有钼铁固氮酶。钒是很少被限制,但钼和铁分别在陆地和海洋环境中是罕见的,因此钼和铁可以分别在相应的生态系统中限制固氮作用。
在固氮作用中,一个电子载体比如铁氧化还原蛋白会首先会还原铁蛋白,随后会还原催化剂组分,这需要铁和催化蛋白的分离和再结合。每固定一分子的氮分子,需要消耗16分子的ATP。额外的生物能量损耗是由于强大的还原剂如铁氧化还原酶的生产和不稳定含氧固氮酶的保护。由于氧气的暴露会使固氮酶失效,故含氧的光能利用菌,比如Trichodesmium spp.、Crocosphaera watsonii 和Nodularia spp. 会使固氮作用和光合作用从空间上(如专门的固氮异形细胞里)或时间上分离。而非光和作用的生物体生活在含氧环境中则需要一些机制从氧气中保护它们的固氮酶,比如增加氧气呼吸、超氧化物歧化酶的解毒作用和固氮酶的构象等机制。而关于存在能够用一个不寻常的固氮酶去进行一个完全不同的、对氧气不敏感的固氮途径的观点也在最近被反驳了。
虽然固氮作用的真核生物还没有被发现,但许多有固氮作用的微生物都可以和真核生物共生。例如单核蓝藻菌UCYN-A是固氮微生物中最普遍的一种,它与小型的单核海藻共生,并在海洋固氮作用中起着重要的作用。共生固氮微生物也是动物肠道菌群的一部分,比如在白蚁类和双壳类中发现的一些特殊细菌。此外,根瘤菌目的一些固氮成员生活在豆科农作物的根瘤中,像苜蓿、黄豆、豌豆和大豆等植物,这些农作物提供了全球20%的食物蛋白。
氨氧化成羟胺
Ammonia oxidation to hydroxylamine
所有已知的好氧氨氧化细菌和古菌都通过使用氨单加氧酶(ammonia monooxygenase, AMO)将氨氧化成羟胺而激活氨。大多数氨氧化细菌属于Betaproteobacteria和Gammaproteobacteria类,并且是属于氨氧化成亚硝酸盐的化学自养生物。它们几乎可以在所有环境中找到,包括施肥土壤和污水处理厂。属于奇古菌门(Thaumarchaeota)的古菌,例如泉古菌(Nitrosopumilus maritimus),也可以通过将氨氧化成亚硝酸盐来进行化学自养的生长。他们的发现解决了海洋中似乎罕见的氨氧化剂的长期谜团。在一些沙质和粉质粘土中奇古菌门比细菌更丰富。此外,嗜酸性氨氧化古菌’Candidatus Nitrosotalea devanaterra’的分离推翻了由于氨的可用性低导致化学自养型氨氧化不能在低pH下发生的普遍假设。许多氨氧化剂,如Nitrosospira sp.和Nitrososphaera viennensis也可以降解有机氮化合物,例如,通过用尿素酶水解尿素以产生额外的氨。古细菌Nitrososphaera gargensis也可以通过用氰化酶水解氰酸盐来产生氨。
最近,在Nitrospira属的成员中也发现了氧化氨的能力,以前认为它只能够进行亚硝酸盐氧化。发现这些将氨氧化成硝酸盐的细菌(完全氨氧化comammox),驳斥了氨和亚硝酸盐氧化需要两组不同微生物的教条。执行完全氨氧化过程的细菌,例如’Candidatus Nitrospira inopinata’似乎很好地适应氨限制环境,并且在氨利用上可以胜过大多数可培养的氨氧化微生物。在氨上生长的完全氨氧化培养物中亚硝酸盐的瞬时积累表明它们比亚硝酸盐更有效地氧化氨。我们假设进行完全氨氧化过程的细菌在氨限制条件下将氨氧化成硝酸盐,并在氧限制条件下进行部分氨氧化成亚硝酸盐。
AMO与甲烷单加氧酶(MMO)密切相关,甲基单加氧酶存在于甲烷氧化菌如γ-变形菌门和候选门NC10(参考文献9)中。尽管效率非常低,MMO也可以将氨氧化成羟胺(图1)。同样,AMO也可以氧化甲烷,但效率低于MMO。有趣的是, (如在地下水中)检测到amo细菌序列执行完全氨氧化过程,但是被错误地指定为丝状甲烷氧化菌Crenothrix polyspora的MMO(pmo)基因微粒。最近对C.polyspora和其他Crenothrix物种的重新测序表明它们实际上含有典型的γ-蛋白质细菌pmo而不是amo。
羟胺氧化成一氧化氮和亚硝酸盐
Hydroxylamine oxidation to nitric oxide and further to nitrite
氨气向羟胺的有氧氧化是一个吸热反应,因此,所有的氨氧化剂可以使羟胺氧化从而保存能量。而需氧氨氧化细菌用八面体血红素羟胺氧化还原酶(HAO)可以将羟胺氧化成亚硝酸盐。但最近研究表明,HAO将羟胺氧化的结果不是亚硝酸盐,而是一氧化氮,然后一氧化氮在一种未知的酶作用下才会被进一步氧化成亚硝酸盐。尽管目前这种未知的催化酶还没有被最终确定下来,但是含铜的亚硝酸还原酶(Cu-NIR,nirk)却可以在一氧化氮被氧化成亚硝酸盐的相反方向起到催化作用。所有的氨氧化细菌,包括新发现的可以一步使氨气彻底氧化成硝酸盐的硝化螺菌属细菌(Nitrospira spp.),它们都包含氨单加氧酶(AMO)和羟胺还原酶(HAO)。相比之下,已知的氨氧化古生菌却不能编码羟胺还原酶(HAO),而使羟胺氧化反应发生的古生菌酶还是未知的。
羟胺氧化还原酶(HAO)属于八面体血红素(octahaem)蛋白家族,在很多微生物中都已经被发现(图2b)。厌氧氨氧化细菌的基因可以编码约十种的类似于HAO的蛋白,而其中的一种也可以使羟胺氧化成一氧化氮。在厌氧氨氧化细菌中,这种羟胺氧化酶(HOX)可以回收从联氨合成酶(HZS)中泄漏的羟胺。
甲烷氧化细菌也可以产生羟胺,这是由于它们非特异的氨氧化反应。属于变形菌门、疣微菌门和NC10菌门中的多种多样的甲烷氧化菌可以编码类似HAO的蛋白,而这种蛋白很有可能使羟胺氧化成一氧化氮,一氧化氮还可以进一步被氧化成亚硝酸盐或者被还原成氧化亚氮( 一氧化二氮/笑气)。目前,在甲烷氧化细菌中发生的这种反应是否可以直接有助于能量保存还未知。
亚硝酸盐向硝酸盐的氧化
Nitrite oxidation to nitrate
亚硝酸盐的氧化是产生硝酸盐的主要生化途径,而这一过程可以通过亚硝酸盐氧化还原酶(NXR)来完成。NXR可以被需氧的亚硝酸盐氧化细菌(包括Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, Chloroflexi, Nitrospinae和 Nitrospirae)、不产氧的光能利用菌(比如荚硫菌属Thiocapsa的KS1、红假单胞菌属Rhodopseudomonas的LQ17)、厌氧氨氧化细菌编码。而需氧的亚硝酸盐细菌在亚硝酸盐氧化还原酶(NXR)的作用下会直接发生亚硝酸盐氧化反应从而来保存能量,但是厌氧亚硝酸细菌却不能做到。此外,厌氧氨氧化细菌会将厌氧亚硝酸氧化反应和碳固定作用结合起来。
亚硝酸氧化细菌具有丰富多样的代谢功能,而且还可以在不是亚硝酸盐的基质上生长。的确,硝化螺菌属的一些细菌可以将氨气彻底氧化成硝酸盐,当硝化螺菌属以硝酸盐为基质进行呼吸作用时,在含氢气环境中生长会进行有氧呼吸,而在有机酸环境中生长则进行厌氧呼吸。亚硝酸盐氧化还原酶(NXR)跟细菌和古生菌的硝酸盐还原酶也相关,亚硝酸盐氧化细菌在NXR的催化作用下进行硝酸盐还原反应。
亚硝酸盐和氨氧化微生物在农业土壤中可以协同作用将氨基化肥转变成硝酸盐,它们在肥料向河水和地下水中的流失过程中起着关键的作用,而化肥的流失会导致河流、湖泊、海岸水域的水体富营养化。与上述同样的协同作用在污水处理工厂中也会被用做去除氮的第一步处理(BOX 2)。在海洋环境中,亚硝酸盐氧化细菌会生产硝酸盐,而硝酸盐是海洋中最主要的生物可利用氮,同时亚硝酸盐氧化细菌也有助于碳的固定。
硝酸盐还原为亚硝酸盐
Nitrate reduction to nitrite
硝酸盐被还原成亚硝酸盐是用来进行呼吸作用的(异化硝酸盐还原反应/dissimilatory nitrate reduction),同时也使氮被同化成生物量。硝酸盐被还原为亚硝酸盐的反应都可以由来自三个领域中的微生物来完成,而这些微生物都存在于含有硝酸盐的缺氧环境中,包括土壤、低氧区、海洋沉积物和人类胃肠道中。这些反应可以被膜结合硝酸还原酶(NAR)或者周质的硝酸还原酶(NAP)催化,而且许多生物体,包括副球菌属的反硝化细菌都含有NAR和NAP。NAR在细胞质中还原硝酸盐,并将质子释放到细胞质中(图3c),并通过质子的动能量直接有助于节约能量。相比之下,NAP在细胞外周质中将硝酸盐还原为亚硝酸盐,因此不改变质子位置。
异化的硝酸盐还原为亚硝酸盐不仅仅是反硝化作用的第一步。许多微生物,比如广泛分布在淡水和海洋沉积物中的巨大硫氧化贝日阿托氏菌就是先将硝酸盐还原为亚硝酸盐,再进一步还原为氨盐。还有许多微生物,比如海洋进化分支SAR11的一些成员只把硝酸盐还原为亚硝酸盐。硝酸盐的还原对于其它氮循环过程来说是亚硝酸盐的一个主要的来源,比如需氧的亚硝酸盐氧化和厌氧氨氧化作用。异化的硝酸盐还原反应还可以与有机物、甲烷、硫化物、氢气或者铁等电子供体的氧化作用相结合。
==硝酸盐是真核生物的主要氮源==,而细菌和古生菌中都含有同化硝酸盐还原酶(NAS)。考虑到硝酸盐支持了至少20%的海洋藻类的生长,所以在海洋中,硝酸盐的同化吸收作用很可能已经超过了氮循环过程中其它大部分的氧化还原反应(BOX 1)。NAS与同化亚硝酸盐还原酶共同产生了氨气,而这氨气也成为了生物量的一部分。因为NAS定位于细胞质中,所以硝酸盐的同化需要用依赖ATP的转运蛋白将硝酸盐转运到细胞内。由于这种能量的需求,NAS的活性会在充满氨的环境中被抑制(比如肥沃的土壤中)。
细菌和古生菌的NAS、周质的硝酸还原酶(NAP)、膜结合硝酸还原酶(NAR)和亚硝酸盐氧化还原酶(NXR)都属于二甲亚砜还原酶家族,而真核生物的同化硝酸盐还原酶属于亚硫酸盐氧化酶家族,这就表明硝酸盐还原酶有多种起源。同化硝酸盐反应与异化硝酸盐还原反应的途径间的差别并不是绝对的,从原理上讲,亚硝酸盐的产生是由于在呼吸链上可以进一步减少硝酸盐的还原反应。相反的是,结核分支杆菌(Mycobacterium
tuberculosis)已被证实可以用NAS复合体来进行硝酸盐的同化吸收。
亚硝酸盐还原为铵盐
Nitrite reduction to ammonium
亚硝酸盐还原为氨盐的反应可以用做同化作用,也可以用作异化作用。异化亚硝酸盐反应大部分是由嗜热的热球菌属(thermophilic Pyrolobus fumarii)家族的细菌来完成的,它们属于Crenarcheota门的甲烷氧化古菌。这种反应是由nrfAH编码的周质的细胞色素c亚硝酸盐还原酶(ccNIR)、八面体血红素亚硝酸盐还原酶(ONR)或者八面体血红素连四硫酸盐还原酶(OTR)催化完成。目前尚不清楚后两种酶是否能用于呼吸作用、亚硝酸盐或羟胺的解毒作用中。亚硝酸盐还原为铵盐的过程中还涉及到中间产物羟胺的形成,而羟胺一直和相应的酶结合在一起直到转变成铵盐。
有趣的是,厌氧氨氧化细菌K. stuttgartiensis在缺少产生铵盐的亚硝酸盐还原酶的情况下,还可以将亚硝酸盐还原为铵盐,因此可以推测亚硝酸盐还原为铵盐是由一种类HAO的蛋白完成的。最近研究表明,一种羟胺还原酶(HAO)可以被ε-变形菌门(比如胎儿弯曲杆菌Campylobacter fetus和鹦鹉螺菌Nautilia profundicola)编码,此酶被称为εHAO,也可以将亚硝酸盐或羟胺还原为铵盐,只不过效率比较低。
异化的亚硝酸盐还原为铵盐的反应是异化硝酸盐还原为铵盐(DNRA)过程的关键反应。微生物可以利用异化硝酸盐还原为铵盐反应(DNRA)和电子供体的氧化耦合来生长,比如有机物、二价铁、氢气、硫化物、甲烷等电子供体。但DNRA对环境的重要性还未可知,而且在海洋和湖泊等沉积物中当与硝酸盐有关的电子供体超量时,DNRA反应就更喜欢反硝化作用。
同化亚硝酸盐还原酶可以产生氨,而且它和NAS一样普遍,这两种酶经常被相同的nas操纵子编码。在海洋中亚硝酸盐最大值的形成是由于浮游植物中硝酸盐的同化吸收作用与亚硝酸盐的还原作用并没有偶联,从而可以释放更多的亚硝酸盐,但这种未偶联的生理原因并不清楚。
亚硝酸盐还原为一氧化氮
Nitrite reduction to nitric oxide
许多微生物都有将亚硝酸盐还原为一氧化氮的能力,比如变形菌、厌氧氨氧化菌和拟杆菌。这些微生物在很多环境中都能被发现,比如在土壤、低氧区、海洋沉积物等环境中(在这些环境中硝酸盐可被利用且含氧量低)。这种反应可以由两种不相关的酶催化,包含血红素的cd1亚硝酸盐还原酶(cd1-NIR; 被nirS编码)和含铜的亚硝酸盐还原酶(Cu-NIR; 被nirK编码),而且这两种酶在细菌和古生菌中都普遍分布。这两种酶都存在于外周质细胞内,不会直接有助于节约能量。这两种酶也可以一起出现在同一个微生物内,比如海洋红嗜热盐菌(Rhodothermus marinus)。
==通常nirS 和nirK 被用作基因标记来研究环境中的反硝化菌==,而且这些基因在其他很多微生物中都存在,包括厌氧氨氧化细菌、亚硝化细菌、甲烷氧化细菌、氨氧化细菌和古生菌。除了cd1-NIR和Cu-NIR,其他亚硝酸盐还原酶也可能存在。比如,一些厌氧氨氧化细菌并不包含nirS 和nirK 这两种基因,但却可以将亚硝酸盐还原为一氧化氮。而为了执行这种反应,这些细菌可以利用类似于HAO的八面体血红素氧化还原酶。
一氧化氮还原为氧化亚氮或氮气
Nitric oxide reduction to nitrous oxide or dinitrogen gas
一氧化氮是一种信号分子,一种有毒物质,也是反硝化作用、硝化作用和厌氧氨氧化作用过程中的一个中间产物。此外,可催化需氧的反硝化作用的细菌可以将2分子的NO变成1分子的N2和1分子的O2。此后,能够使NO还原的微生物在环境中被广泛发现,包括废水处理工厂、农业土壤、海洋沉积物、海洋低氧区。由微生物引起的NO还原反应是N2O的主要来源,而N2O是一个强大的温室气体(比CO2的温室效应强烈310倍,也是破坏臭氧层的主要物质。能够产生N2O的一氧化氮还原酶(NOR)被用于一氧化氮的解毒作用或呼吸作用。属于不同组的酶从黄素蛋白到血红素铜氧化酶,它们都通过生命树而广泛分布。黄铜-二铁蛋白例如黄铜-二铁一氧化氮还原酶(NORvw)可以通过像硫酸盐还原细菌Desulfovibrio
gigas来解毒一氧化氮。其他的NOR类型的酶是从真菌线粒体中发现的依赖NADH细胞色素P450的一氧化氮还原酶,如镰刀菌属的马拉色菌Fusarium oxysporum以及最近在大肠杆菌Escherichia coli中发现的混合聚类蛋白(hybrid cluster protein, HCP)。
血红素铜氧化酶家族包括末端氧化酶类,比如依赖红细胞色素cNOR、依赖对苯二酚qNOR和含铜的依赖对苯二酚CuANOR,它们都在利用NO呼吸中起作用。氧化亚氮是反硝化作用的中间产物,而NOR存在于反硝化细菌P. denitrificans和假单胞菌属Pseudomonas stutzeri等微生物中。氧化亚氮对于一些微生物来说也可以是最终产物,比如绿叶假单胞菌Pseudomonas chlororaphis。氨氧化细菌在硝化-反硝化作用中可以产生氧化亚氮,NOR是被用来在亚硝酸盐还原过程中还原产生的NO的。在氨氧化细菌和能够一步彻底氧化氨到硝酸盐的可用细菌中,氧化亚氮也可以通过细胞外的中间产物一氧化氮和羟胺的非生物反应产生。此外,氨氧化细菌还可以在羟胺氧化中产生的一氧化氮还原酶(NOR)的催化作用下产生氧化亚氮。和氨氧化细菌相似,嗜甲烷细菌也可以通过在羟胺氧化和亚硝酸盐还原中NOR的催化作用产生氧化亚氮。与此相比,氨氧化古生菌只是通过中间产物NO和羟胺的非生物反应产生氧化亚氮。
氮肥的使用大幅度增加了氧化亚氮的排放。由于氮转化微生物的协同作用,3%-5%的氮作为农业氮肥被转化成氧化亚氮。氮肥越来越多的被用来种植农作物,用于生物燃料生产,这为代替化石燃料提供了可能。但也因此形成了一个困境,越多的氮肥被用来生产生物燃料,就会有越多的氧化亚氮被排放出来。为了生产生物燃料而对氮肥的使用,抵消了由于减少对化石燃料的使用降低的温室效应的效果。
NO转变成氮气和氧气的歧化作用是最近被发现的新的氮转化反应(图1)。从湖泊和湿地等富含甲烷和硝酸盐的缺氧系统中发现的微生物(比如’Ca. Methylomirabilis oxyfera’)用这种歧化反应从亚硝酸盐中产生自己的氧气。’Ca. Methylomirabilis oxyfera’可以在缺氧环境中生存,也可以利用需氧的甲烷氧化途径。这种歧化反应包含一种不寻常的酶qNOR,暂时称这种酶为一氧化氮歧化酶(NO-D)。一氧化氮的歧化反应比之前预想的更为普遍,与此相似的是,这种不寻常的酶qNOR的基因序列在其它菌门中也存在,比如γ-变形菌门的HdN1菌株和拟杆菌门的Muricauda ruestringensis。
氧化亚氮还原为氮气
Nitrous oxide reduction to nitrogen gas
由微生物引起的氧化亚氮向氮气的还原,是减少这种强大温室气体的主要途径。已知的催化这种反应的酶只有氧化亚氮还原酶(nitrous oxide
reductase, NOS),而这种酶定位于细胞外周质中,这对于由质子动能引起的能量节约并没有直接的促进作用。来自于变形菌门、拟杆菌们、绿菌门的多种多样的细菌与来自于泉古菌门、嗜盐菌门的古生菌都能使用NOS。在大肠杆菌中发现的稍微不同的能够编码NOS的基因揭示了人们忽视了土壤中NOS基因序列的多样性。编码NOS变体的生物经常不含有其它NO还原酶。一些真核生物,比如有孔虫类Foraminifera和网足虫Gromiida,目也可以减少氧化亚氮,但是它们的酶学机制未知。
长久以来,人们都认为NOS比其他氮化合物还原酶对氧气、pH、硫化物更敏感。基于这种明显的敏感性,氧化亚氮向环境中的排放完全归因于在能够使硝酸盐被还原为氮气的完全反硝化菌生物体内NOS的活性被抑制。此外,比如土壤和海洋环境,由于这些不完全反硝化菌的相作,比只能进行亚硝酸盐还原为氧化亚氮反应或者化亚氮还原为氮气的反应的微生物生态位的差别也会导致氧化亚氮的产生和消耗间的不平衡。
联氨合成和氧化为氮气
Hydrazine synthesis and hydrazine oxidation to dinitrogen gas
人们普遍认为只有氧气才能激活氨气,而生物可利用氮的损失只有通过反硝化作用被转化为氮气。然而最近以亚硝酸盐作为终端电子受体转化成氮气的厌氧氨氧化反应却推翻了这两种教条观点。HZS是唯一已知的可以激活厌氧氨的酶,而且它只在浮霉菌门中的五种厌氧氨氧化细菌中被发现。HZS也是唯一已知的可以使两种离散的氮化合物形成N-N化合键的酶,在一个含有两步的反应中会产生中间产物联氨。而联氨合成的假设机制是从一氧化氮还原为羟胺开始的(图2d),然后羟胺和氨一起发生归中反应产生联氨,而联氨在自然界中是最强有力的还原剂之一。在这种反应中,羟胺从一个活动位点转移到另一个活动位点(图2d),这样在联氨合成酶HZS的作用下消耗羟胺。编码HZS的基因有两个,分别是hzsA和hzsB,这两种基因一般会被作为环境中厌氧氨氧化细菌的标记基因。
联氨脱氢酶(hydrazine dehydrogenase, HDH)可以将联氨氧化成氮气。通过对氨基酸的测序,知道了这种酶与羟胺氧化还原酶(HAO)和羟胺氧化酶(HOX)有关,但被羟胺抑制且只氧化联氨。联氨的氧化作用发生在被称为厌氧氨氧化体的一种独特的膜结构中,而且很可能与节省能量有直接关系。有趣的是,厌氧氨氧化细菌的所有分解代谢酶(HDH, HZS, NIR, HOX 和 NXR)都只存在于厌氧氨氧化体中。
HDH主要负责向大气中释放氮气。在上一个10年,关于在海洋中的厌氧氨氧化过程已经很清楚了,而且该过程在陆地生态系统中也有着非常重要的作用。
Box 2 | 利用微生物去除废水的氮
Box 2 | Nitrogen removal by microorganisms in wastewater treatment
自工业革命以来,农耕、化石燃料的燃烧、家庭和工业产生的废水等一直都是造成氮污染的主要因素,而氮污染严重影响着地球上的生命。在过去的四十年,当发现过量的氮会导致水体富营养化以及氨盐的毒性可以造成鱼类的死亡时,氮就被认为是废水中重要的污染物,因此,在原本用来去除有机碳的废水处理工厂里都又添加了氮去除系统,然而大部分常规的废水处理工厂并没有进行对氮的去除。
在大多数自然生态系统中,珍贵的氮都是是被循环回收和保存的,但与此对比,在废水处理厂却需要通过特殊设计将水体中的铵盐转变成氮气释放到大气中。在废水处理厂中,有机碳会首先被去除,然后将有机碳贫乏但铵盐氮丰富的水体送入到氮去除系统中。传统的去氮系统是依靠硝化作用(NH4+ → NO2- → NO3-)将氨盐氧化成硝酸盐,再依靠反硝化作用(NO3- → NO2- → NO → N2O → N2)将硝酸盐还原为氮气。但硝化作用需要大量曝气增加氧气来创造条件使氨盐氧化成硝酸盐(平均每1分子的氨需要2分子的O2),然后通过外源添加有机碳(一般为甲醇)诱导将硝酸盐还原为氮气的反硝化作用。因此传统的去氮方法很费财力、精力和资源,而且这个过程中还会产生温室气体N2O。为了缓和这个问题,现在很多不同结构的反应器应运而生,如将富含有机碳的部分原始废水直接输入到反硝化作用的步骤中,或者对单独的一个池子里进行间接性的曝气,以促进硝化作用和反硝化作用的进行。
在过去数十年里,厌氧氨氧化的出现,被用作去除氮的可供选择的一种方法。在一些简洁的生物反应器中,需氧的氨氧化细菌(如亚硝化单胞菌属Nitrosomonas europea)在氧气限制条件下可以将一半的生物可利用氨转化为亚硝酸盐,称为部分亚硝化。接下来在细菌‘Candidatus Kuenenia stuttgartiensis’作用下发生厌氧氨氧化,将亚硝酸盐和剩下的氨盐转变成氮气(图BOX 2中实线所示)。在这种部分亚硝化厌氧氨氧化的系统中,可以生成硝酸盐的需氧亚硝酸盐氧化剂(比如硝化螺菌属Nitrospira spp. 或者硝化杆菌属 Nitrobacter spp.)是不受欢迎的,因为它们降低了氮去除的效率。这种亚硝化厌氧氨氧化生物反应器与传统的氮去除系统相比,需要的曝气较少,而且不需要再另添加有机碳,产生的N2O气体也较少。目前,这种亚硝化厌氧氨氧化生物反应器在污水处理(比如来自厌氧污泥硝化器的废水)中也越来越多的被应用,而这些系统在含有较低铵盐浓度的全市规模的废水处理中的应用,为可持续的治理废水铺平了道路。
最近发现的一些氮循环中的微生物也可以应用到废水治理中。可以使氨氧化到亚硝酸盐的古生菌和彻底氨氧化为硝酸盐的细菌在废水处理工厂中被发现,但它们在这些系统中所扮演的角色还很不清楚。在氧气受限制的氮去除系统中(比如亚硝化厌氧氨氧化反应器),那些可以进行彻底的氨氧化成硝酸盐的细菌可能跟传统的氨氧化过程一样产生亚硝酸盐。而令人兴奋的是,新发现的依赖亚硝酸盐和依赖硝酸盐的厌氧甲烷氧化微生物为废水的治理提供了一种新的可能。一个联合厌氧甲烷氧化菌(比如‘Candidatus Methylomirabilis’
spp.和‘Candidatus Methanoperedens’ spp.)联和可进行厌氧氨氧化反应的微生物反应器,就可以同时去除氨盐、硝酸盐和甲烷(图BOX 2中虚线所示)。这样的系统已经在实验室的条件下建立起来了,但这种完整规模的污水处理系统还没有在实际环境中被应用,而在这样系统中,需氧的甲烷氧化剂如Methylomonas spp.可以有助于甲烷的去除。
对氮循环微生物的基本生理和生化以及应的研究正在步步进行中,新的微生物的发现会引领更高效和持续发展的去氮系统,反之亦然。很明显地,这种趋势可以保护环境而造福我们的子孙后代。
Box 2 | 利用微生物去除废水的氮
微生物构成的氮转化网络
微生物在氮转化方面的多样性是惊人的,每一种微生物都有各自最优的生长条件。由于在自然界中生长环境是多变的,而且很少能达到最优条件,所以微生物个体对氮的转化效率必然是低效的。然而,在自然界中依靠微生物群落的氮转化效率比依靠单个微生物的转化效率高多了。因此,在微生物群的作用下,几乎没有生物可利用氮逃逸到大气中,而以氮气的形式损失的少量的氮也会由于固氮作用而达到平衡。这种明显的氮体内平衡不仅是全球生物圈的特征,也是很多生态系统的特征,比如森林土壤和海洋环流。这些能够高效进行氮回收的必需微生物群落,即使因为环境扰动而使微生物的物种组成发生了改变,它们仍然会保持氮转化的反应。由微生物连接起来的氮转化过程在自然和人为建造的生态系统中都形成了一个复杂的网络(图3)。
世界上最大的生态系统-海洋环流,因为大量的氮回收利用而达到了一个氮平衡(图3a)。在这里主要的氮转化过程是由蓝藻细菌如Prochlorococcus marinus完成氮的同化作用、由浮游动物和异养细菌如‘Candidatus Pelagibacter ubique’完成的氨化作用和由Nitrosopumilus spp. 和Nitrospina spp. 完成的硝化作用(BOX 1,图3a)。而由微生物如Trichodesmium spp.和‘Ca. Atelocyanobacterium’完成的固氮作用,它们在海洋环流中属于相当小的氮转化过程。但因为可发生固氮作用的区域非常大,所以它是向海洋中提供生物可利用氮的主要途径。
与海洋环流相比,那些低氧区的水域虽然不到公海海域的1%,但却占海洋领域氮损失的30%-50%(BOX 1)。在这里,厌氧微生物如‘Candidatus Scalindua’ spp.可以与需氧的生物体如Nitrosopumilus spp. 和 Nitrospina spp. 共生。由微生物构成的氮转化网络在公海的低氧区是非常复杂的,彼此之间以及与旁边发生的氮转化过程都是已知的(图3b)。
和低氧区水域相似,在废水处理工厂也是以不平衡的氮转化为特点。这些人为设计的系统主要用来将铵盐转变成氮气排到空气中的(BOX 2)。
农业土壤是人为建造的最大的生态系统,它们由微生物构成的氮转化系统已经受到了人工输入氮的强烈影响。种植的豆科农作物和固氮微生物群形成了共生关系,这大大增加了环境中氮的输入量。固氮微生物菌如Bradyrhizobium spp.)经常生活在特定的根瘤中,并给豆科植物提供铵盐(图3c),而铵盐泄漏到周围土壤中又可以促进其他微生物的氮转化过程,比如需氧的氨氧化作用。在稻田领域,人工肥料的使用导致了强烈的硝化作用,并加大了氮的损失。最近的研究展示 高度复杂的氮转化网络,包括亚硝酸盐还原‘Ca. Methylomirabilis’ spp.和硝酸盐还原(‘Ca.
Methanoperedens’ spp.)甲烷氧化菌 (图3d)。
在这些生态系统中,一些氮转化微生物,如厌氧氨氧化菌,可以进行多重氧化还原反应(反应1、2、5、7、10、12和13;图1)。但是,硝化和反硝化等过程是由一个复杂的特异网络以模块化的方式执行的(图3)。这种模块化是氮转化微生物网络的一个普遍特征,导致了合作和竞争的相互作用(如图3所示)。亚硝基槐属Nitrosopumilus spp.和硝化菌属Nitrospina spp.之间存在协同作用,它们共同将氨氧化成硝酸盐(图3a,b)。在大多数环境中,硝化是由氨氧化微生物和亚硝酸盐氧化微生物的不同组合进行的,它们也分别与氨和亚硝酸盐竞争。具有非常不同代谢的微生物之间也存在着底物竞争,如硝化螺属Nitrospira spp.、‘Candidatus Brocadia’ spp.、‘Ca. Methanoperedens’ spp.和假单胞菌属,它们都在竞争亚硝酸盐(图3d)。微生物间的相互作用也可以同时合作和竞争:亚硝基硫菌Nitrosopumilus spp. 为‘Ca Scalindua’spp.生产亚硝酸盐,但两者都竞争氨(图3b)。
控制这些相互作用的因素不太清楚。有时,单一的生理特性被用来解释某些氮转化微生物在环境中的优势。例如,在氨耗尽的环境中,氨氧化古细菌相对于细菌的丰度是由于古细菌具有较高的氨亲和力。然而,最近有研究表明,陆生细菌“嗜氨芽孢杆菌”‘Ca. Nitrospira inopinata’的氨亲和力高于所有培养的陆生氨氧化古细菌。然而,执行Comammox过程的微生物并不能主导所有氨耗尽的陆地环境。氮转化微生物的成功还取决于其他因素,如使用替代底物和细胞能量需求。这些因素在形成氮转化微生物网络中可能具有一般作用。
图3 | 不同生态系统的氮转换网络
Fig. 3 | Potential nitrogen-transforming microbial networks in different ecosystems
*图3a. 公共海水环流氮循环网络图
a | 公海环流营养有限地区进行广泛氮循环。在阳光照射的地表水中,蓝藻主要吸收铵和/或有机氮化合物进行生长。浮游动物分解的病毒裂解释放出有机氮如尿素由异养细菌矿化成铵。固氮细菌提供额外的铵。在深水中,铵被氧化成硝酸盐。其中一些硝酸盐扩散到地表水中,被浮游植物吸收。
a | The open ocean gyres are vast nutrient-limited regions in which nitrogen is extensively recycled. In the sunlit surface waters, cyanobacteria mainly assimilate ammonium and/or organic nitrogen compounds for growth. Viral lysis and grazing by mesozooplankton release organic nitrogen (for example, urea), which is subsequently mineralized back to ammonium by heterotrophic bacteria. Nitrogen-fixing bacteria provide additional ammonium. In deeper waters, ammonium is oxidized to nitrate. Some of this nitrate diffuses up into the surface waters and is assimilated by phytoplankton.
图3b. 海水低氧区氮循环网络
b | 海洋氧气最低区位于海洋东部边界,在那里,风驱动的富营养水上升流刺激地表水的初级生产力。随后下沉有机物的好氧矿化会耗尽底层水中的氧气。好氧硝化群落能很好地适应低氧条件,对亚硝酸盐和硝酸盐进行氨氧化。由于厌氧氨氧化菌的活性和较小程度的反硝化作用,最低氧区是氮流失的主要区域。反硝化过程涉及复杂的微生物群落。
b | Marine oxygen minimum zones are found on the eastern boundaries of oceans, where wind-driven upwelling of nutrient rich waters stimulates primary productivity in the surface waters. The subsequent aerobic mineralization of sinking organic matters depletes oxygen in the underlying waters. Aerobic nitrifying communities that are well adapted to low oxygen conditions perform ammonia oxidation to nitrite and nitrate. The oxygen minimum zones are major regions of nitrogen loss owing to the activity of anaerobic ammonium-oxidizing bacteria and to a lesser extent denitrification. Complex communities of microorganisms are involved in the denitrification process.
图3c. 人造最大生态系统——农田氮循环网络
c | 最大的人造生态系统是用于农作物生产的农田。豆类是常见的作物,也是蛋白质的重要来源。它们通过释放有机碳来影响周围土壤中的微生物群落,并与固氮微生物共生,如慢生根瘤菌。漏入周围土壤中的铵可以促进好氧氨和亚硝酸盐氧化。随后硝酸盐在土壤中向缺氧区扩散,为氮转化过程提供燃料,如硝酸盐还原成铵、一氧化二氮和氮气。
c | Among the largest man-made ecosystems are agricultural fields that are used for crop production. Legumes are common crops and an important source of protein. They influence the microbial community in the surrounding soil by releasing organic carbon and live in symbiosis with nitrogen-fixing microorganisms, such as Bradyrhizobium spp. Ammonium that leaks out into the surrounding soil can fuel aerobic ammonia and nitrite oxidation. Subsequent diffusion of nitrate to anoxic zones in soil fuels nitrogen-transforming processes, such as dissimilatory nitrate reduction to ammonium, nitrous oxide and dinitrogen gas.
图3d. 稻田的氮循环网络
d | 稻田是被水淹没的农田,用尿素等含氮化合物进行施肥以种植水稻。尿素水解和固氮产生氨,氨在水稻植物根系周围的氧化土壤中被氧化成硝酸盐。硝酸盐随后扩散到潜在的缺氧土壤燃料过程中,如反硝化、厌氧氨氧化(anammox)和产甲烷产生的甲烷氧化。
d | Rice paddies are flooded agricultural fields, which are fertilized with nitrogen-containing compounds such as urea to grow rice155. Urea hydrolysis and nitrogen fixation generate ammonia, which is oxidized to nitrate in oxic soils surrounding the rice-plant roots. Subsequent diffusion of nitrate to the underlying anoxic soil fuels processes such as denitrification, anaerobic ammonium oxidation (anammox) and the oxidation of methane produced by methanogenesis.
Box3 | 未被发现的生物化学反应
在过去十年里许多由微生物引起的新的氮转化反应和途径被发现,而基于热力学方面的考虑,应该会存在由微生物引起的更进一步释放能力的反应(反应15—26)。然而这里有一些反应可以由已知的酶催化,但有一些反应却需要迄今为止未被发现的生物化学催化剂(反应15—19,25和26)。例如依赖硝酸盐的氨氧化作用(反应15)不能通过厌氧氨氧化途径来进行,因为氨盐需要首先被氧化成中间产物羟胺或者类似的含氧物质。
反应 15:5NH4+ + 3NO3- → 4N2 + 9H2O + 2H+ (ΔG0ʹ = −297 kJ mol-1 NH4+)
相似的,激活氨盐的新的生化途径对于依赖铁和锰的氨氧化作用是必需的(反应16—19)。
反应 16:NH4+ + 6Fe3+ +2H2O → NO2- + 6Fe2+ + 8H+ (ΔG0ʹ = −247 kJ mol-1 NH4+)
反应 17:2NH4+ + 6Fe3+ → N2 + 6Fe2+ + 8H+ (ΔG0ʹ = −303 kJ mol-1 NH4+)
反应 18:NH4+ + 3MnO2 + 4H+ → N2- + 3Mn2+ + 4H2O (ΔG0ʹ = −60 kJ mol-1 NH4+)
反应 19:2NH4+ + 3MnO2 + 4H+ → N2 + 3Mn2+ + 6H2O (ΔG0ʹ = −249 kJ mol-1 NH4+)
相反地,一些歧化反应(反应20—24)可以由已知的微生物用现存的生化机制实施。厌氧氨氧化细菌可以进行对亚硝酸盐(反应20)和一氧化氮(反应21)的歧化作用。
反应 20:5NO2- + 2H+ → N2 + 3NO3- + 2H2O (ΔG0ʹ = −60 kJ mol-1 NO2-)
反应 21:10NO + 2H2O → 3N2 + 4NO3- + 4H+ (ΔG0ʹ = −100 kJ mol-1 NO)
相似的,由亚硝酸盐转化成氧化亚氮和硝酸盐的歧化反应(反应22)、由一氧化氮转化成亚硝酸盐和氧化亚氮的歧化反应(反应23)或者是由一氧化氮转化成硝酸盐和氧化亚氮的歧化反应(反应24),在理论上可以由需氧的亚硝酸盐氧化细菌完成。
反应 22:4NO2- + 2H+ → N2O + 2NO3- +H2O (ΔG0ʹ = −32 kJ mol-1 NO2-)
反应 23:4NO + H2O → 2NO2- + N2O + 2H+ (ΔG0ʹ = −40 kJ mol-1 NO)
反应 24:8NO + H2O → 2NO3- + 3N2O + 2H+ (ΔG0ʹ = −56 kJ mol-1 NO)
这些微生物可以利用亚硝酸盐氧化还原酶使亚硝酸盐氧化成硝酸盐,而存在于硝化杆菌属Nitrobacter spp.、硝化球菌属Nitrococcus spp.、硝化螺菌属Nitrospira spp.和硝化刺菌属Nitrospina spp.中的亚硝酸盐还原酶可以将亚硝酸盐还原为一氧化氮。一氧化氮的氧化可以在硝化杆菌属Nitrobacter spp.中观察到,但是这种反应是生物反应还是非生物反应还未可知,而且与该反应有关的酶还不清楚。一氧化氮氧化成亚硝酸盐的反应可以由含铜的亚硝酸盐还原酶(由nirk基因编码)催化,众所周知这个反应是双向的。NO还原为N2O的反应可以由末端氧化酶类催化,而末端氧化酶类在进化上是与NO还原酶有关。
N2O是一种强效的温室气体,在缺氧条件下可以被还原为氮气,而它在好氧条件下被认为在生物学上性质比较稳定的。有趣的是,从热力学上讲需氧的氧化亚氮被氧化成亚硝酸盐或者硝酸盐反应是可行的(反应25、26),但这种反应需要新的生化途径。
反应 25:N2O + O2 +H2O → 2NO2- + 2H+ (ΔG0ʹ = −21 kJ/mol N2O)
反应 26:N2O + 2O2 + H2O → 2NO3- + 2H+ (ΔG0ʹ = −89 kJ/mol N2O)
将催化这些未被发现反应的微生物鉴定出来的唯一方法就是在条件可控的实验室内培养它们。很显然,对这些氮转化微生物的生理生化条件的研究在未来几年的里是研究热点。
结束语
Concluding remarks
确定形成氮转化网络的因素需要对所涉及微生物的生理、生态和进化有更深入的了解。所有微生物中只有一小部分被培养,未被培养的大多数可能含有未被发现的代谢途径(BOX 3)。培养再加上严格的生化、生理和基因组特征,已经改变了我们对关键氮循环过程的认识。好氧亚硝酸盐氧化细菌和厌氧氨氧化细菌具有迄今为止未预料到的代谢多样性,这使得它们的分类仅仅是好氧亚硝酸盐氧化剂或厌氧氨氧化剂的分类方式严重不足。许多好氧亚硝酸盐氧化剂可能在环境中以氢氧化剂、氨氧化剂或硝酸盐还原剂的形式生长。厌氧氨氧化菌也可以使用短链脂肪酸、甲胺和铁(II)作为电子受体,它们可以使用硝酸盐、锰(IV)和铁(III)作为电子受体。
相反,越来越多的人认识到单一微生物的完全反硝化是例外,而不是普遍规律,许多微生物是只进行一种或几种氮氧化物还原反应的专家。专门的氮氧化物还原剂往往缺乏已知的基因,使他们能够将硝酸盐一路还原到氮气。这些专业的氮氧化物还原剂通常被描述为不完整的反硝化剂,这与将氨氧化剂(如亚硝酸盐)描述为不完整的硝化剂类似。
毫无疑问,按照经典的六氮循环过程对生物体进行分类将变得越来越困难,这就让观察者不得不确定生物体的功能。如果我们能从过去几十年的研究中学到一件事,那就是微生物间没有严格的界限。为了生存,他们会做任何必要的事情。
参考文献
Kuypers M. M. M., Marchant H. K. & Kartal B. The microbial nitrogen-cycling network. Nat Rev Microbiol. 2018, 16: 263-276. doi: https://doi.org/10.1038/nrmicro.2018.9
综述 | Nature Reviews Microbiology:微生物构成的氮循环网络(IF:26.819)
猜你喜欢
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature Cell专刊 肠道指挥大脑
系列教程:微生物组入门 Biostar 微生物组 宏基因组
专业技能:学术图表 高分文章 生信宝典 不可或缺的人
一文读懂:宏基因组 寄生虫益处 进化树
必备技能:提问 搜索 Endnote
文献阅读 热心肠 SemanticScholar Geenmedical
扩增子分析:图表解读 分析流程 统计绘图
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
在线工具:16S预测培养基 生信绘图
科研经验:云笔记 云协作 公众号
编程模板: Shell R Perl
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难,我们建立了“宏基因组”专业讨论群,目前己有国内外5000+ 一线科研人员加入。参与讨论,获得专业解答,欢迎分享此文至朋友圈,并扫码加主编好友带你入群,务必备注“姓名-单位-研究方向-职称/年级”。PI请明示身份,另有海内外微生物相关PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
学习16S扩增子、宏基因组科研思路和分析实战,关注“宏基因组”
点击阅读原文,跳转最新文章目录阅读